An Extreme Value Theory Approach to Wildlife Disease Transmission
Francisca Javiera Rudolph, PhD
University of Florida George A. Smathers Libraries
One Health Center of Excellence
Averages Hide the Story
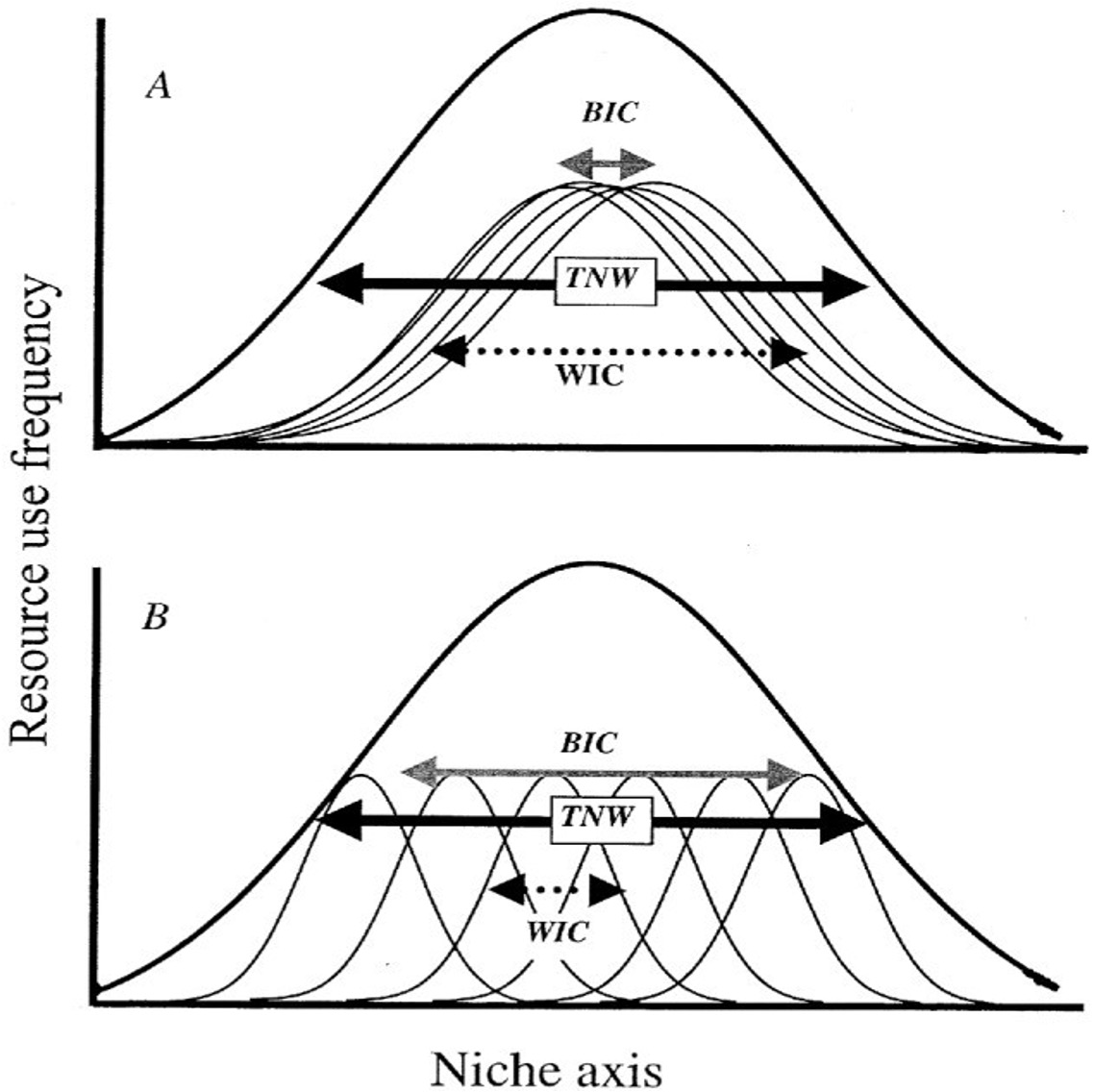
Bolnick and colleagues looked across 93 species and found that individual specialization is widespread across taxa - different individuals are doing different things
Focusing on variation among individuals, not species averages, reveals the mechanisms behind seed dispersal outcomes.
Bolnick et al. The American Naturalist 2003
Zwolak. Biological Reviews 2018
The Origin: Aracari & Seed Dispersal
Primary dispersers of Virola trees in Ecuadorian rainforest
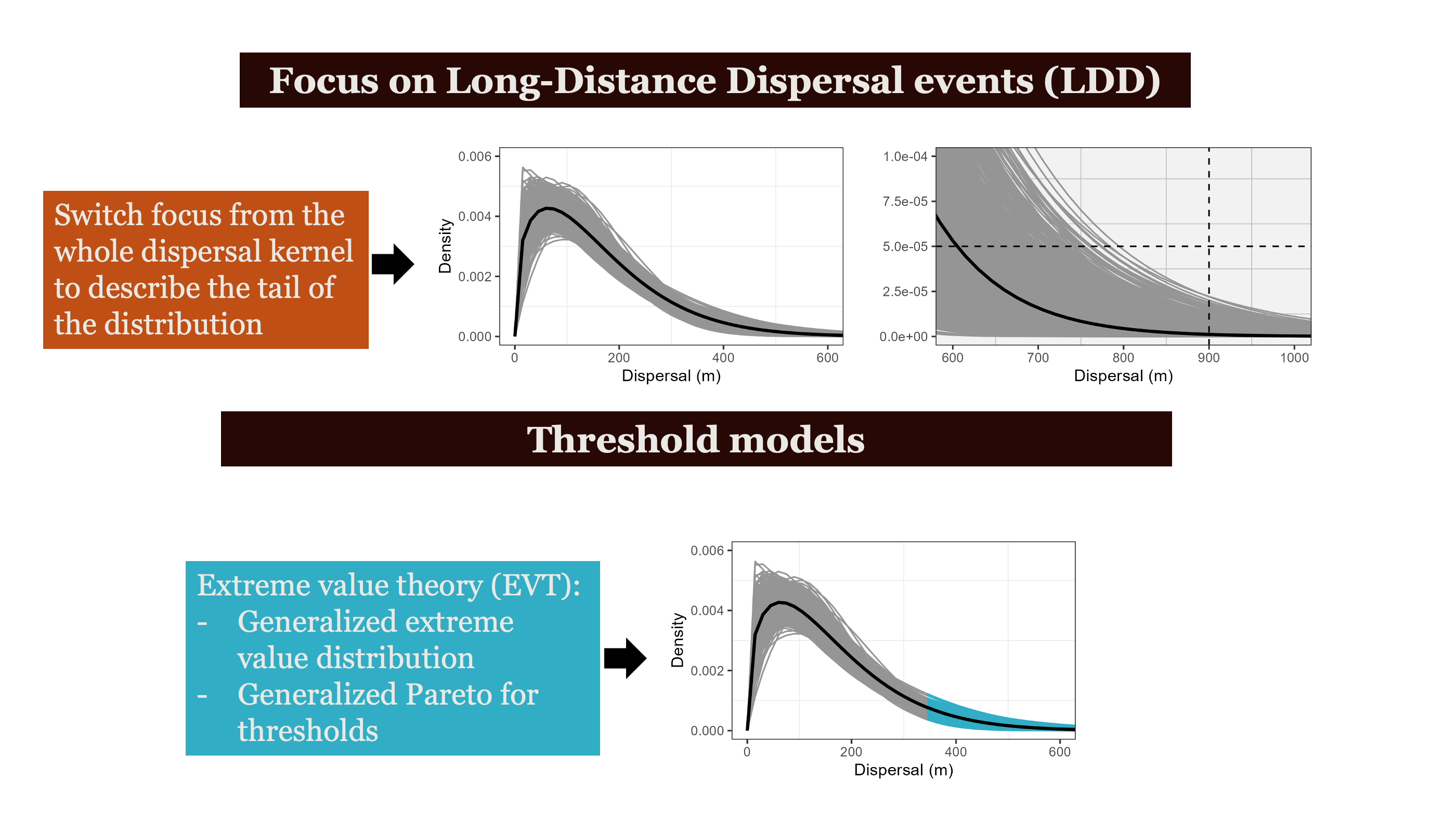
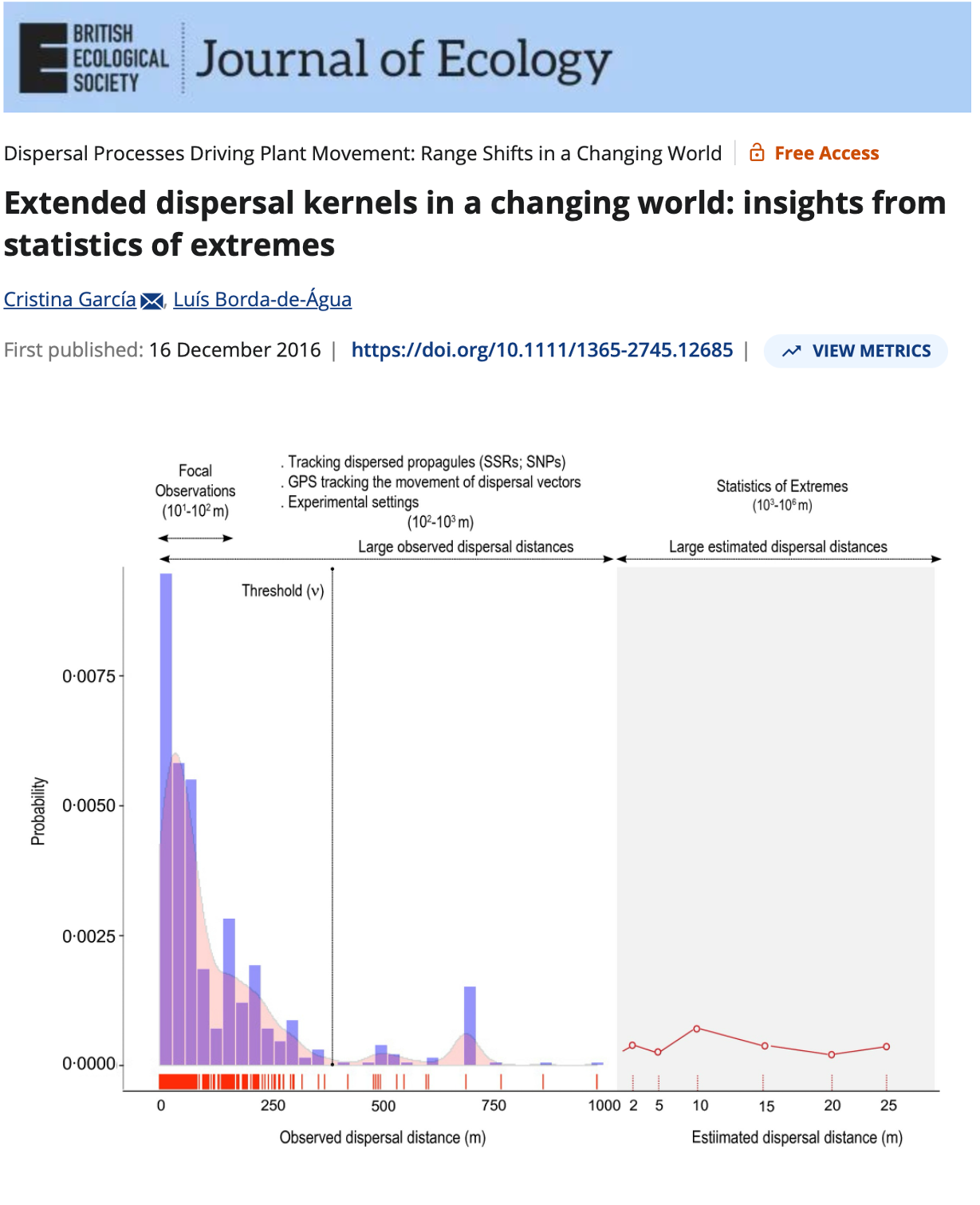
Most seeds land close to parent — but rare long-distance events are not that rare when we account for individual variation in movement
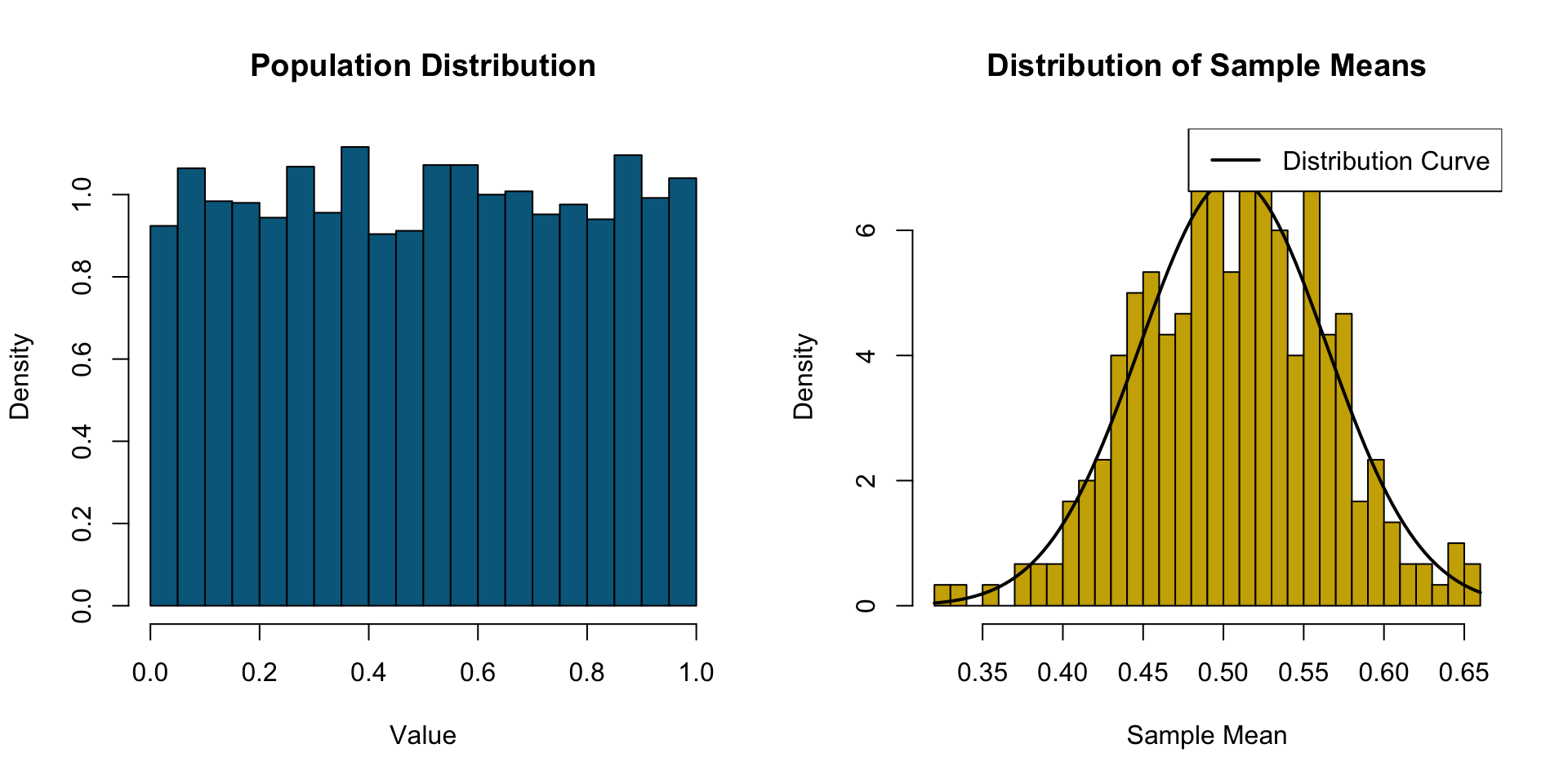
Central Limit Theorem (CLT) & Extreme Value Theory (EVT)
The special rules in statistics
CLT = gather data -> take average -> repeat many times
averages are normally distributed
The Central Limit theorem states that the distribution of sample means approaches a normal distribution as the sample size increases, regardless of the population’s original distribution.
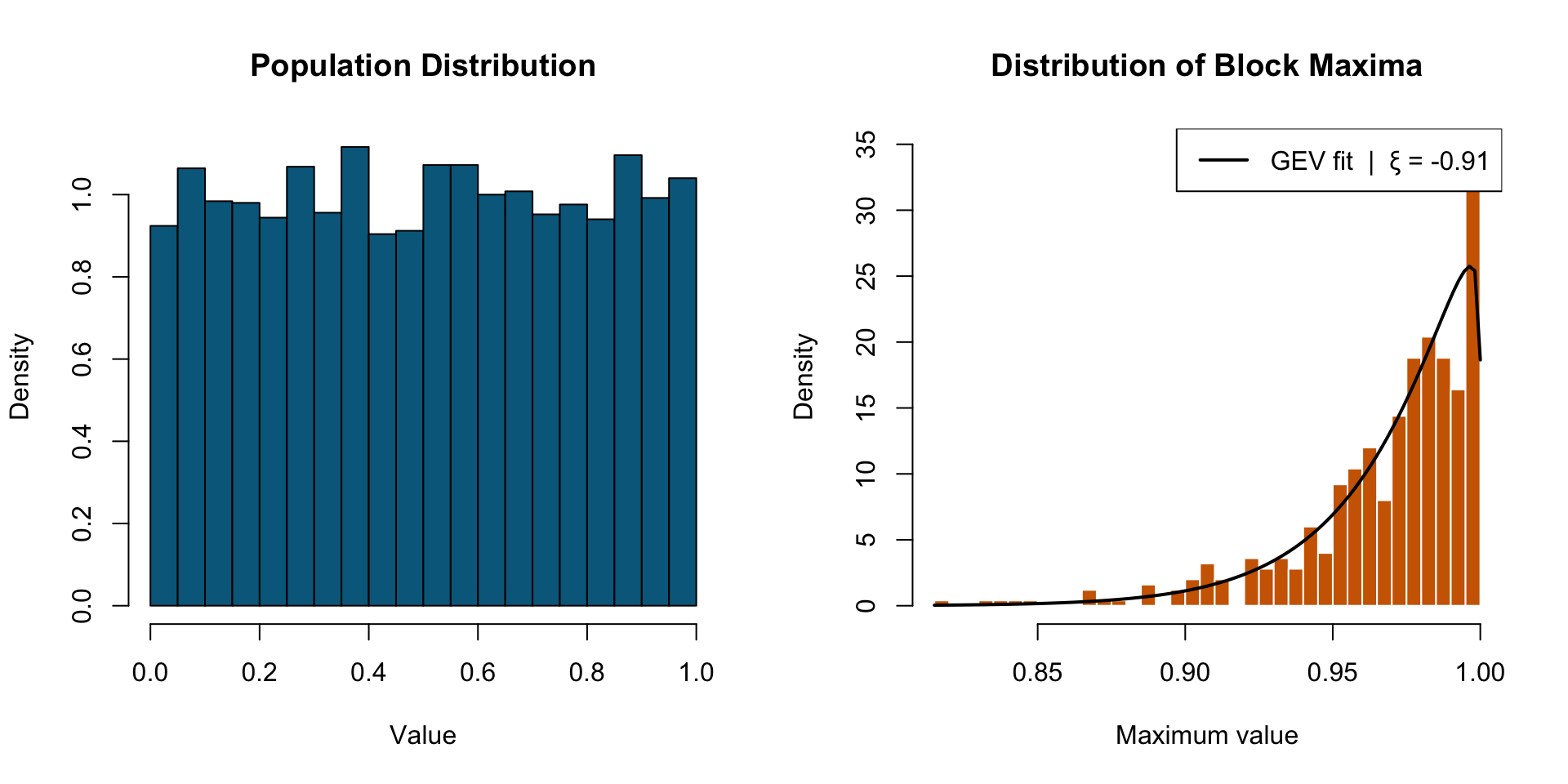
Extreme Value Theory states that regardless of the underlying data-generating process, the behavior of extreme observations — the rare, large events in the tail — converges to a Generalized Extreme Value distribution. The tail has its own universal structure.
EVT
From Pattern to Process
Individual variation in movement shapes the tail of the dispersal kernel —
and we can use ξ (shape) to see this
The process
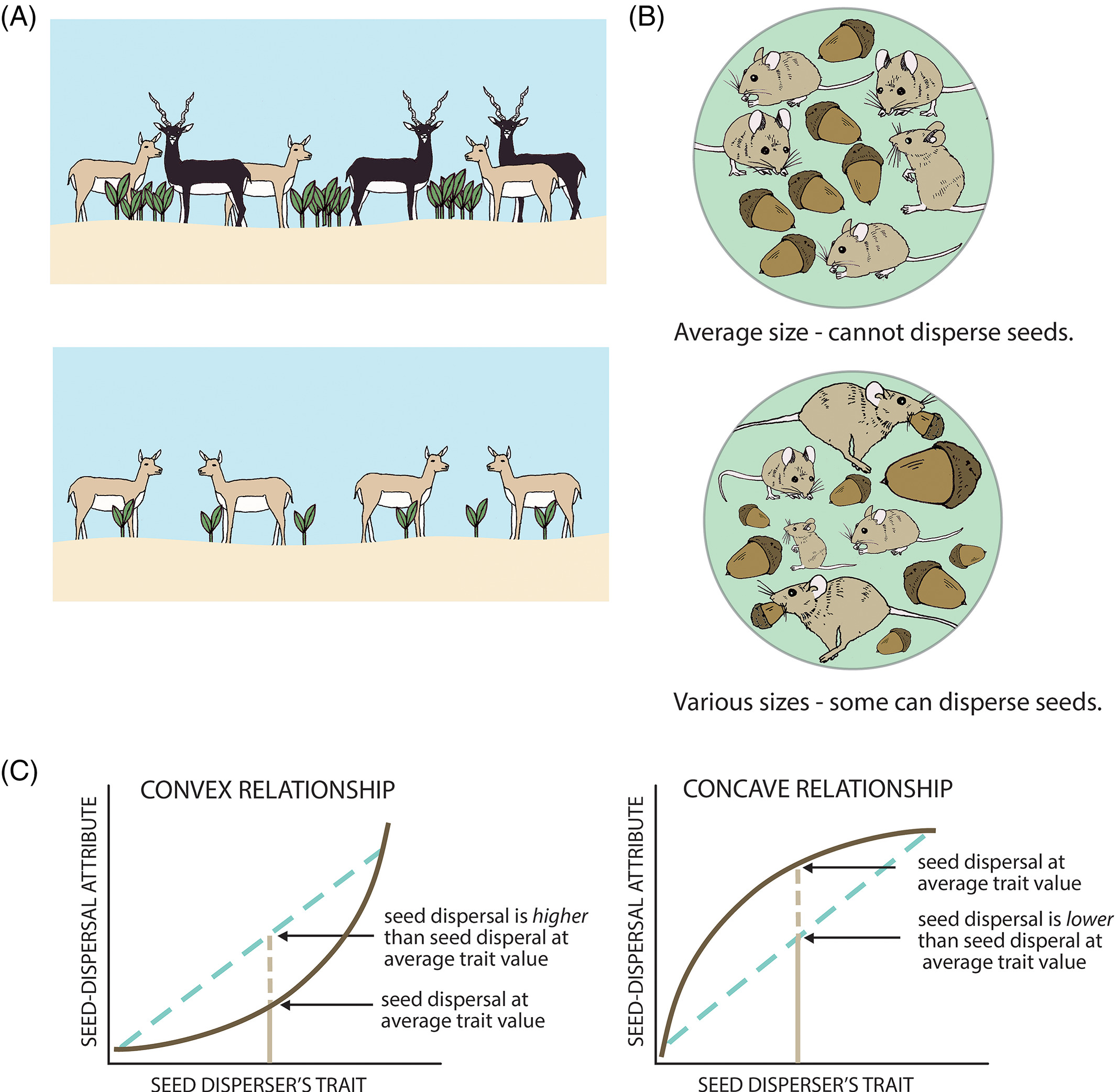
Individual frugivores differ in how far they move. Some are homebodies. Some are explorers. That variation is not just noise, it is the mechanism.
The pattern
Pooled models underestimate long-distance dispersal. When individual variation is modeled explicitly, the tail of the seed shadow is fatter — and ξ > 0 confirms it.
The link
ξ is not a fitting artifact. It is a fingerprint of the movement process. The shape of the seed dispersal tail traces back to how individuals move and how we capture variation across individuals.
EVT has offered ecologists a framework for ‘rare’ events like long-distance dispersal since the 1990s — the tools were there before the field was ready to use them.
Gaines& Denny (1993). The largest, smallest, highest, lowest, longest, and shortest: extremes in ecology. Ecology.
Katz et.al. (2005). Statistics of extremes: modeling ecological disturbances. Ecology
Beisel et.al (2007). Testing the extreme value domain of attraction for distributions of beneficial fitness effects. Genetics
Individual Variation in Disease Transmission
“Population-level analyses often use average quantities to describe heterogeneous systems…”
— Lloyd-Smith et al. Nature 2005
Sound familiar?
\(R_0\) is an average. It hides the same individual variation we saw in dispersal kernels:
Most individuals infect zero or one
A few infect many — superspreaders
Individual reproductive number, \(\nu\), as a random variable representing the expected number of secondary cases caused by a particular infected individual.
Same \(R_0\), six different epidemiological worlds.
More heterogeneity means more outbreaks die out, but the ones that don’t are explosive.
k describes the shape of that individual variation
The Mechanistic Foundation
Ponciano & Capistrán (2011) derive the incidence rate from first principles:
An infected individual has realized disease dispersion\(a\)
Number of successful transmission encounters \(X(a)\) follows a pure birth process
Individual variation matters here:
The exponential distribution for \(\Lambda\) already allows individuals to differ in how much they disperse pathogen.
But the exponential has a thin tail — it does not generate superspreaders.
What if dispersion effort follows a heavier-tailed distribution? What if — like seed dispersers — some infected individuals are true long-distance movers?
From Movement to Superspreaders: The Lomax Emerges
What if dispersion effort \(a\) is itself heterogeneous?
Allow dispersion effort \(a\), to be distributed as an exponential distribution, whose parameter is sampled from a gamma distribu- tion:
Let \(A \sim \text{Gamma}(\theta, \tau)\) — individual variation in transmission potential — and \(Y \mid A \sim \text{Exp}(A)\) — realized dispersion given that potential.
Then the marginal distribution of \(Y\) is:
\[f_Y(y) = \int_0^\infty a e^{-ay} \cdot \frac{\theta^\tau}{\Gamma(\tau)} a^{\tau-1} e^{-\theta a} \, da = \frac{\tau \theta^\tau}{(y + \theta)^{\tau+1}}\]
This is the Lomax distribution — special case of a GPD - a heavy-tailed distribution with support on \([0, \infty)\).
The tail emerges mechanistically from the compound structure.
And the probability of transmission under Lomax-distributed dispersion:
\(\theta\) and \(\tau\) are not abstract parameters — they characterize how individuals in a population move. Some animals are homebodies, some are explorers.
That variation across individuals, captured by the Gamma, is what drives the heavy tail.
Allow the movement to vary, and EVT drops out the other end.